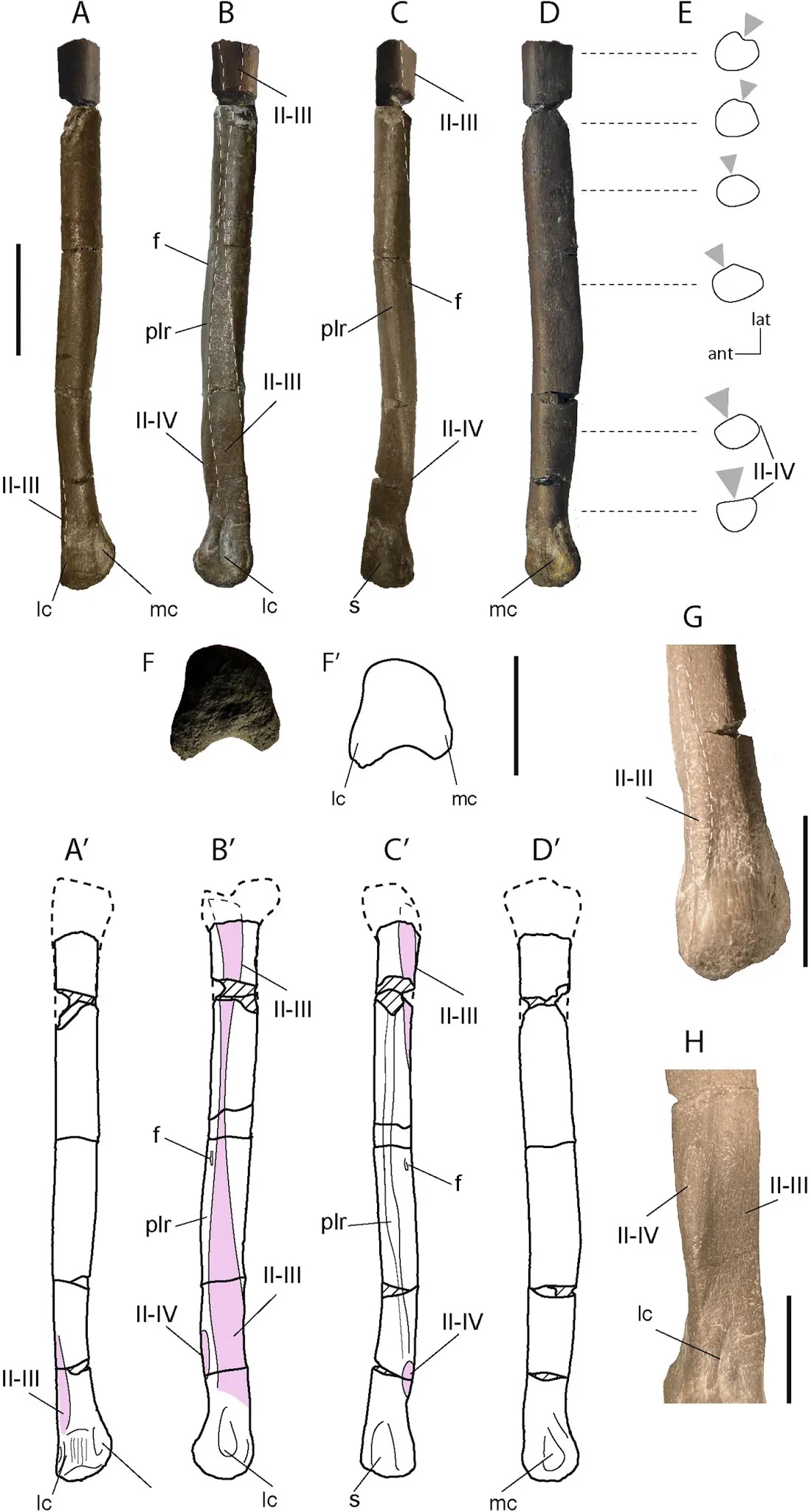
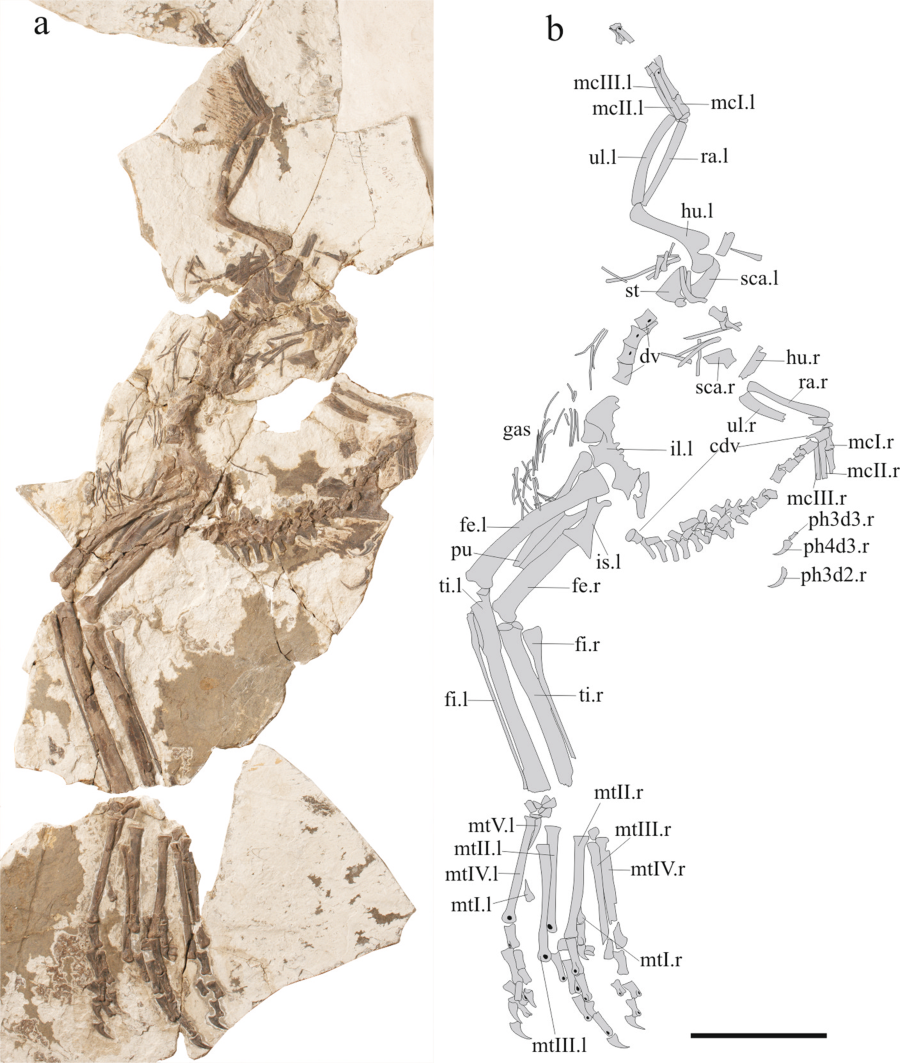
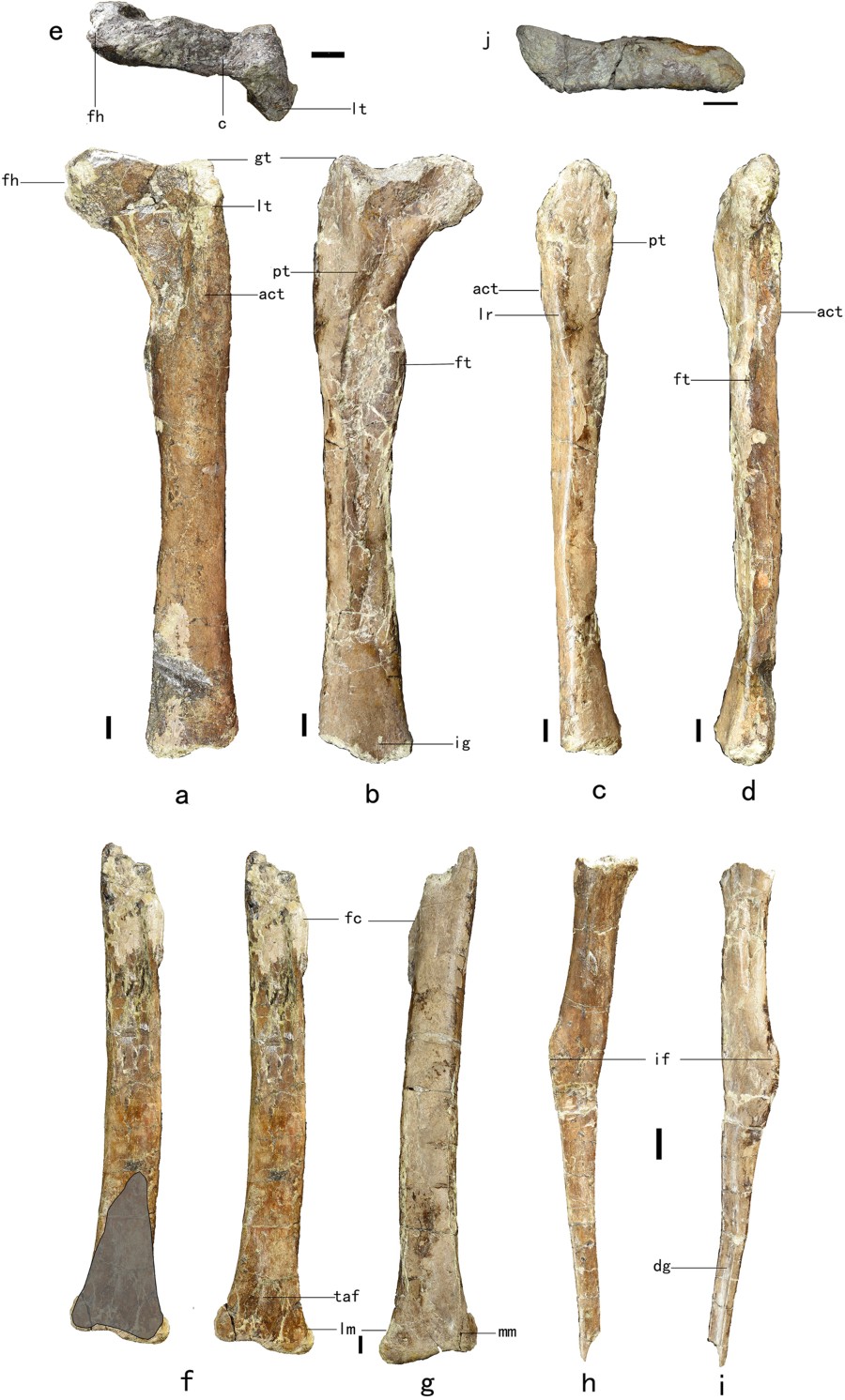
Nel genere
cinematografico poliziesco e d'azione, esiste una coppia
stereotipata, la così detta coppia "poliziotto buono - sbirro
cattivo". Entrambi i membri della coppia sono "dei buoni"
ed entrambi combattono il crimine, ma risultano antitetici da un
punto di vista "aristotelico". Il poliziotto buono è
generalmente "formale", vuole rispettare le regole sempre e
comunque, ed è perfettamente aderente al regolamento. Egli incarna
l'idea che le regole siano da rispettare sempre e comunque, costi
quel che costi. Lo sbirro cattivo è invece allergico alle
regole, non rispetta il regolamento e tende a fare "a modo suo".
Egli incarna l'idea che la giustizia e l'etica sono più importanti
della forma, e che il nobile fine di far trionfare la giustizia
richiede ai buoni, a volte, di "agire da cattivi".
Generalmente, la simpatia dello spettatore va verso "lo sbirro
cattivo", se non altro perché spesso la trama di questi film è
proprio incentrata sulla necessità di "sporcarsi le mani"
e "violare il regolamento" per combattere il crimine. Una
scena classica di questi film è appunto il momento in cui lo sbirro
cattivo spiega all'amico poliziotto buono che, a volte, occorre
violare il regolamento proprio per mantenere in vita il senso stesso
di quel regolamento. Perché, dopo tutto, le regole sono fatte per
essere infrante. e la forma ha senso solamente in funzione della
sostanza.
Vi ricordate il brutto
affare che coinvolse la revisione tassonomica dell'oviraptoride
"
Ingenia"
yanshini? Ne parlai alcuni anni fa,
quando scoprii (e
denunciai
su questo blog) che un certo Easter, autore della "revisione"
di
Ingenia (revisionato con il nome di
Ajacingenia),
aveva scritto quell'articolo
plagiando
il blog di Mickey Mortimer e non contattando Rinchen Barsbold,
paleontologo mongolo di fama internazionale, ancora in vita ed autore
della specie originaria "I."
yanshini,
quindi non coinvolgendolo nella revisione della specie istituita
proprio da Barsbold nel 1981.
Formalmente, il nome
"Ajacingenia" è pienamente valido, perché rispetta
tutte le regole del Codice Internazionale di Nomenclatura Zoologica.
Secondo l'approccio del "poliziotto buono", il nome
Ajacingenia chiude la faccenda, e deve essere usato. Eppure,
io ho più volte sottolineato che tale nome è, per quanto
"formalmente buono", "sostanzialmente cattivo",
sbagliato ed ingiusto. Pertanto, io non lo ritengo un termine
scientificamente valido, per questioni "etiche" più che
formali: usare quel termine significa avallare un episodio di plagio
e di scorrettezza etica. Oltre che ingiusto nella sostanza, esso
diventa un pericoloso precedente di vandalismo tassonomico e cattiva
condotta professionale.
Altri preferiscono
"rispettare il regolamento" e ignorare la questione etica:
essi dicono che siccome Ajacingenia è formalmente valido, non
ci sono motivi per non usarlo. Io la penso diversamente. Forse perché
io non sono un poliziotto buono, ma uno sbirro cattivo: una persona
poco rispettosa della forma ma molto attenta alla sostanza.
All'inizio di questo
anno, ricevetti la richiesta di fare la revisione tecnica ad un
manoscritto scritto da Greg Funston, Mendonca, Currie e Barsbold,
avente come oggetto una dettagliata revisione anatomica e tassonomica
degli oviraptorosauri dal Bacino del Nemegt. In questo articolo, gli
autori revisiovano la morfologia di "Ingenia" ed
usavano il nome "formalmente valido" di Ajacingenia.
Alla fine della mia revisione, inclusi una nota non vincolante, ma
che mi auguravo che gli autori avrebbero preso in considerazione:
evitare di usare Ajacingenia, per i motivi etici che ho menzionato
prima, e di "aggirare il codice" proponendo la seguente
soluzione:
In tutte le recenti
filogenesi degli oviraptorosauri, "Ingenia/Ajacingenia"
yanshini risulta sister taxon di Heyuannia huangi, un
oviraptoride cinese. I due oviraptoridi sono molto simili, in
particolare nella inusuale morfologia robusta della mano, al punto
che le differenze tra i due olotipi sono a livello di dettagli
minori. Pertanto, proposi, perché non abbandonare Ajacingenia
(per motivi etici sostanziali) e proporre una "espansione"
del nome Heyuannia, così da includere al suo interno non solo
la specie huangi, ma anche la specie yanshini? Ovvero,
proposi a Funston et al. di istituire una nuova combinazione
"Heyuannia yanshini" al posto di "Ajacingenia
yanshini". Dato che il genere Heyuannia è stato
istituito 11 anni prima di Ajacingenia, esso è un potenziale
sinonimo senior di questo ultimo, e quindi ha priorità nel caso i
due nomi si riferiscano al medesimo taxon. Pertanto, "espandendo"
l'inclusività di Heyuannia (non più associato ad una sola
specie, ma a due), si rende inutile la necessità di Ajacingenia.
In questo modo, sicuramente poco formale, ma sostanzialmente valido e
comunque permesso dal Codice, si rimuoveva alla radice il problema
etico generato da Ajacingenia.
Notate che non ci sono
motivi per rifiutare questa opzione: esattamente come Gorgosaurus
libratus e Albertosaurus libratus sono due alternative
valide per il medesimo taxon (dipende da quanto inclusivo
consideriate il nome Albertosaurus), allo stesso modo possiamo
usare Heyuannia yanshini e Ajacingenia yanshini. Con la
differenza, per me sostanziale, che Heyuannia yanshini è
eticamente più accettabile di Ajacingenia yanshini.
 |
| Da Funston et al. (2017) |
Ieri, l'articolo di
Funston et al. (2017) è stato pubblicato. Gli autori hanno accolto
il mio suggerimento e propongono la combinazione Heyuannia
yanshini.
Essi scrivono:
Barsbold
(1981) originally described this oviraptorid as “Ingenia”
yanshini, but the genus was preoccupied by a nematode. Despite
recognition of the taxonomic issue, it was not resolved until Easter
(2013) coined “Ajancingenia”. There are, however, several
ethical problems with the study of Easter (2013), including the
plagiarism of text and a figure. Hence, despite the validity of
‘Ajancingenia’ under the ICZN, we propose synonymizing
Ajancingenia and Heyuannia to avoid an ethical dilemma.
This synonymy is supported by recent oviraptorosaur phylogenies
(Lamanna et al., 2014; Funston and Currie, 2016; Lü et al., 2016),
which find H. yanshini and H. huangi as sister taxa.
Funston mi ha
personalmente risposto ieri, via Facebook, ringraziandomi per
aver proposto una "soluzione elegante" ed eticamente
risolutrice alla controversia tassonomica. Io, ovviamente, seguirò
questa proposta, non tanto perché l'ho proposta io, ma perché mi
pare veramente la soluzione più corretta, nella sostanza, ad una
brutta faccenda di plagio e vandalismo tassonomico.
Bibliografia:
Funston,
G.F. et al. (2017) Oviraptorosaur anatomy, diversity and ecology in
the Nemegt Basin. Palaeogeography, Palaeoclimatology, Palaeoecology
http://dx.doi.org/10.1016/j.palaeo.2017.10.023





.jpg/1920px-Dinosaur_fossil_(1).jpg)









