
 |
| Credit: Eva Hoffman / The University of Texas at Austin |
Lo scorso anno, Hoffman e
Rowe (2018) pubblicarono lo studio su un fossile eccezionale dal
Giurassico Inferiore degli USA: i resti associati di 39 esemplari del
mammaliaforme tritilodontide Kayentatherium.
I
tritilodontidi sono il clade di cinodonti più prossimi
evolutivamente ai primi mammiferi (nota pedante tassonomica che puoi
anche saltare: qui uso “mammifero” nel senso più ampio che
include anche forme come Morganucodon in Mammalia, e non nel
senso stretto del crown group). I tritilodontidi sono sovente citati
negli studi e nei modelli relativi all'origine del piano corporeo del
nostro clade, proprio perché ci mostrano un probabile aspetto della
condizione ancestrale di tutti i mammiferi. Un clade, Mammalia, il
cui nome (non è un dettaglio secondario), ha la propria radice nella
parola “mamma”, per rimarcare l'importanza biologica delle
cure parentali materne basate sull'allattamento.
L'origine
dei mammiferi è più o meno legata, biologicamente ed
adattativamente, all'origine delle cure parentali basate
sull'allattamento? La questione va oltre la semplice analisi
filogenetica deducibile dai fossili, perché tocca un elemento
fisico-fisiologico-psicologico che ha plasmato le nostre peculiari
forme sociali e relazionali: il legame madre e figlio.
Tutti
amano la mamma, quindi, qualsiasi scoperta ci aiuti a capire
l'origine della condizione mammaliana - e della condizione di mamma
mammaliana - non può che suscitare interesse e passione.
Forse
troppa passione.
Almeno,
questa è la mia impressione dopo aver letto lo studio pubblicato in
questi giorni da Benoit (2019) che propone una rivalutazione critica
dello scenario adattativo che Hoffman e Rowe (2018) hanno proposto a
loro volta a seguito della scoperta dell'associazione dei 39
esemplari di Kayentatherium. Rivalutazione che non intacca il
valore della scoperta, ma la libera dall'inutile fardello di
aspettative generali che, più o meno inconsciamente, ognuno di noi
carica su un fossile come questo.
Procediamo
per gradi.
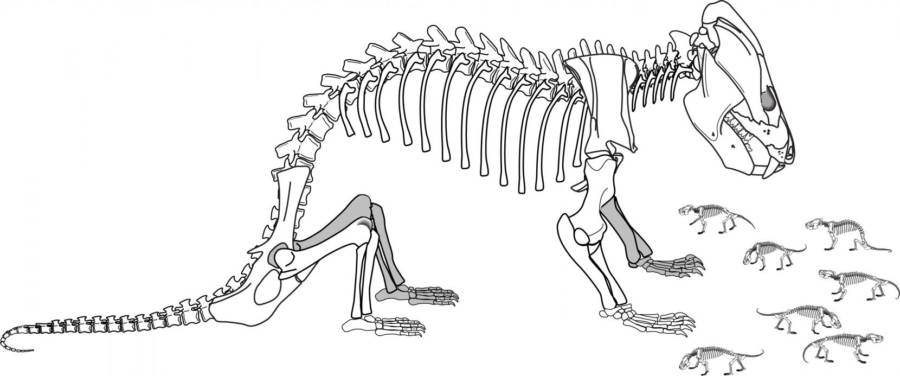
Hoffman
e Rowe (2018) descrivono l'associazione di 39 Kayentatherium.
La particolarità dell'associazione è che dei 39 esemplari, solo uno
è un adulto, mentre tutti gli altri 38 sono dei minuscoli esemplari
molto immaturi. Siccome una così numerosa associazione di piccoli
insieme ad un adulto è stata scoperta in un clade di therapsidi
evolutivamente (e morfologicamente) molto prossimi ai primi
mammiferi, gli autori concludono che questa sia una associazione
familiare: vista la onnipresente relazione madre-cuccioli nei
mammiferi, l'adulto è quindi da considerare la madre dei 38 piccoli.
Quale
scoperta migliore per capire l'origine del clade “mamma”-liano
che scoprire una mamma di “quasi”-mammifero?
(Notate
che nel giro di un paio di righe siamo passati dalla descrizione dei
fatti ad una inferenza filogenetica - impropria, come spiego sotto -
e da questa siamo saltati su una interpretazione comportamentale che
si adatta molto bene alle nostre aspettative generali in merito a
Mammalia)
Ed è
qui che l'analisi di Hoffman e Rowe (2018) diventa intrigante (o,
come va oggi di moda, “groundbreaking”): essi devono
necessariamente concludere che il modus riproduttivo di
Kayentatherium fosse prettamente rettiliano (produzione di
tante piccole uova), dato che nessun mammifero partorisce così tanti
cuccioli alla volta. Quella che gli autori hanno scoperto è una
covata “da lucertola” in un animale che è praticamente una
specie di “nutria” giurassica. Il ragionamento è corretto (ma
non implica che la premessa fosse corretta), e comunque di per sé
non particolarmente rivoluzionario, dato che anche i mammiferi
monotremi attuali (come l'ornitorinco) si riproducono tramite uova
(sebbene non producano covate così numerose!). La condizione ovipara
nei monotremi era già una prova sufficiente per escludere che i
cinodonti permo-triassici avessero evoluto l'ovoviparità o la
viviparità. Quello che però nessuno si aspettava, e che rende
l'interpretazione “groundbreaking”, è stato lo scoprire che
persino i “quasi-mammiferi” come Kayentatherium
producessero “ancora” così tante uova alla volta!
(Notare
i miei virgolettati: qui sto volutamente cavalcando le nostre
aspettative evoluzionistiche, montando una controversia più di
pregiudizi che di fatti. Ma così avviene con fossili eccezionali che
aprono scenari su momenti “topici” della nostra evoluzione)
Un
dettaglio chiave per risolvere la faccenda è nella bocca dei piccoli
di Kayentatherium, i quali non mostrano differenze di
dentizione rispetto all'adulto: ovvero, essi non mostrano i denti “da
latte” bensì la formula dentaria adulta. Siccome la difiodontia
(ovvero, la sostituzione con la crescita da una dentizione “da
latte” ad una permanente) è un carattere chiave dei mammiferi
legato all'allattamento, dobbiamo concludere che Kayentatherium
non allattasse i propri piccoli. E, in effetti, l'idea che una
singola madre possa allattare 38 piccoli alla volta appare
fisicamente, fisiologicamente (e psicologicamente) impossibile.
Pertanto, tutto concorre a escludere una forma di allattamento in
Kayentatherium. E, di conseguenza, anche in tutti i sinapsidi
non-mammiferi, che sono ancora più distanti dai mammiferi rispetto a
Kayentatherium: i loro piccoli erano tutti pienamente in grado
di nutrirsi in autonomia. Dimenticatevi i cuccioli di Dimetrodon
da allattare: erano piuttosto come piccoli alligatori pronti ad
azzannare qualunque cosa a tiro di mandibole.
Benoit
(2019) rivaluta alcuni elementi della faccenda in maniera più fredda
e spassionata rispetto alla narrazione fatta sopra, una freddezza
quasi oltraggiosa verso la mamma ancestrale che morì coi suoi 38
cuccioli nel Giurassico Inferiore.
Benoit
(2019) si domanda, quasi banalmente, se l'associazione dei 38
Kayentatherium possa avere una spiegazione differente rispetto
alle cure parentali. Dopo tutto, non c'è scritto sul fossile che
esso sia una famiglia: quello che vediamo è solo una associazione di
conspecifici. Benoit (2019) fa un rapido censimento di tutte le
associazioni “adulto-immaturi” nel record fossile dei sinapsidi,
e nota che in tutti gli altri casi, spesso relativi a taxa molto più
lontani dai mammiferi rispetto a Kayentatherium, ritroviamo un
numero minore di giovani, ma sempre di dimensioni (rispetto
all'adulto) più grandi rispetto ai piccoli Kayentatherium. Ad
esempio, mentre i 38 Kayentatherium sono grandi solo il 5%
dell'adulto, normalmente si trovano solamente da uno a 3 giovani
associati assieme, e sempre grandi almeno il 30% dell'adulto. Sebbene
il campione sia relativamente basso per fare delle statistiche
robuste, è comunque indicativo che all'aumentare delle dimensioni
dei giovani trovati associati rispetto all'adulto, diminuisce il loro
numero. Che fine facevano gli altri 35 piccoli, se a metà della
crescita ne restavano solo 3? La mamma evidentemente non si struggeva
molto per l'ecatombe di figli a cui assiteva ad ogni nuova nidiata.
Concludiamo, senza particolari soprese, che la mortalità infantile
fosse molto elevata, e che solo pochi esemplari raggiungevano la
dimensione adulta. Anche questo elemento, nota Benoit (2019) va a
sfavore di cure parentali intense “alla mammifero” (e, noto io,
ricorda invece il pattern che osserviamo nei dinosauri).
Come
spiegare allora l'associazione di 38 piccoli se questi non era
allattati dalla madre, se questi erano autonomi nel nutrirsi e se poi
la stessa madre evidentemente non pareva curare loro grandi cure e
difese? In natura, esistono numerosi motivi “non-familiari” per
cui gli animali si aggregano, a volte anche con animali di altre
specie. Proprio di un altro sinapside triassico è l'associazione con
un anfibio, entrambi fossilizzati assieme dentro la medesima cavità.
Evidentemente, questi animali avevano la tendenza a stare nascosti in
tane, e queste tane erano occupate da più animali, anche di specie
diverse. Viste le condizioni climatiche fortemente aride e stagionali
di molti contesti fossili permo-triassici, è probabile che
l'estivazione dentro tane sotterranee fosse una strategia molto
diffusa tra i sinapsidi di dimensioni medio-piccole. Aggregarsi per
affrontare la stagione arida dentro una tana comune è quindi una
strategia opportunistica che non implica necessariamente cure
parentali o allattamento: “pernottare” assieme riduceva il
rischio di essere preda di qualche carnivoro e permetteva di
massimizzare la conservazione del calore corporeo.
Pertanto,
il paradosso dei 38 piccoli Kayentatherium associati ad un
adulto potrebbe avere una soluzione meno romantica e più prosaica
rispetto allo scenario della super-mamma: l'aggregazione
opportunistica di una nidiata di giovani assieme ad un adulto in una
tana comune. Attenzione: questo non esclude che l'adulto fosse la
madre, ma nemmeno lo dimostra: alla luce di tutti gli elementi,
stabilire se l'adulto fosse un genitore non pare nemmeno
fondamentale, dato che, come scritto sopra, i giovani Kayentatherium
non erano allattati e quindi non avevano bisogno di un adulto per
essere nutriti. E prima che possiate tirare in ballo
mamma-Kayentatherium che va a caccia per i piccoli, ricordate
che i tritilodontidi erano erbivori, e che avendo già la dentizione
adulta, ogni esemplare era perfettamente autonomo nel nutrirsi fin
dalla nascita.
Volendo
modernizzare comunque questo fossile, possiamo concludere che “Mamma
Kayentatherium” era solo l'affittuario del tugurio dove
bivaccavano i giovani durante la stagione secca...
Non me
ne vogliano male le mamme in ascolto, ma il fatto che noi mammiferi
abbiamo una tale predilezione per il rapporto mamma-figli non implica
automaticamente che ogni fossile di sinapside da qui fino al
Carbonifero debba essere interpretato come una preparazione del
nostro peculiare e dispendioso sistema riproduttivo e parentale.
Bibliografia:
Benoit
J. 2019. Parental care or opportunism in South African Triassic
cynodonts? South African Journal of Sciences. 115(3/4).
Hoffman
EA, Rowe TB. 2018. Jurassic stem-mammal perinates and the origin of
mammalian reproduction and growth. Nature. 561:104–108.












