
Questo post è da annoverare nella
lista dei 50 post più nerd nella storia della blogosphera.
Tutto è stato ispirato da questa
immagine postata su twitter e Facebook da Mark Witton:
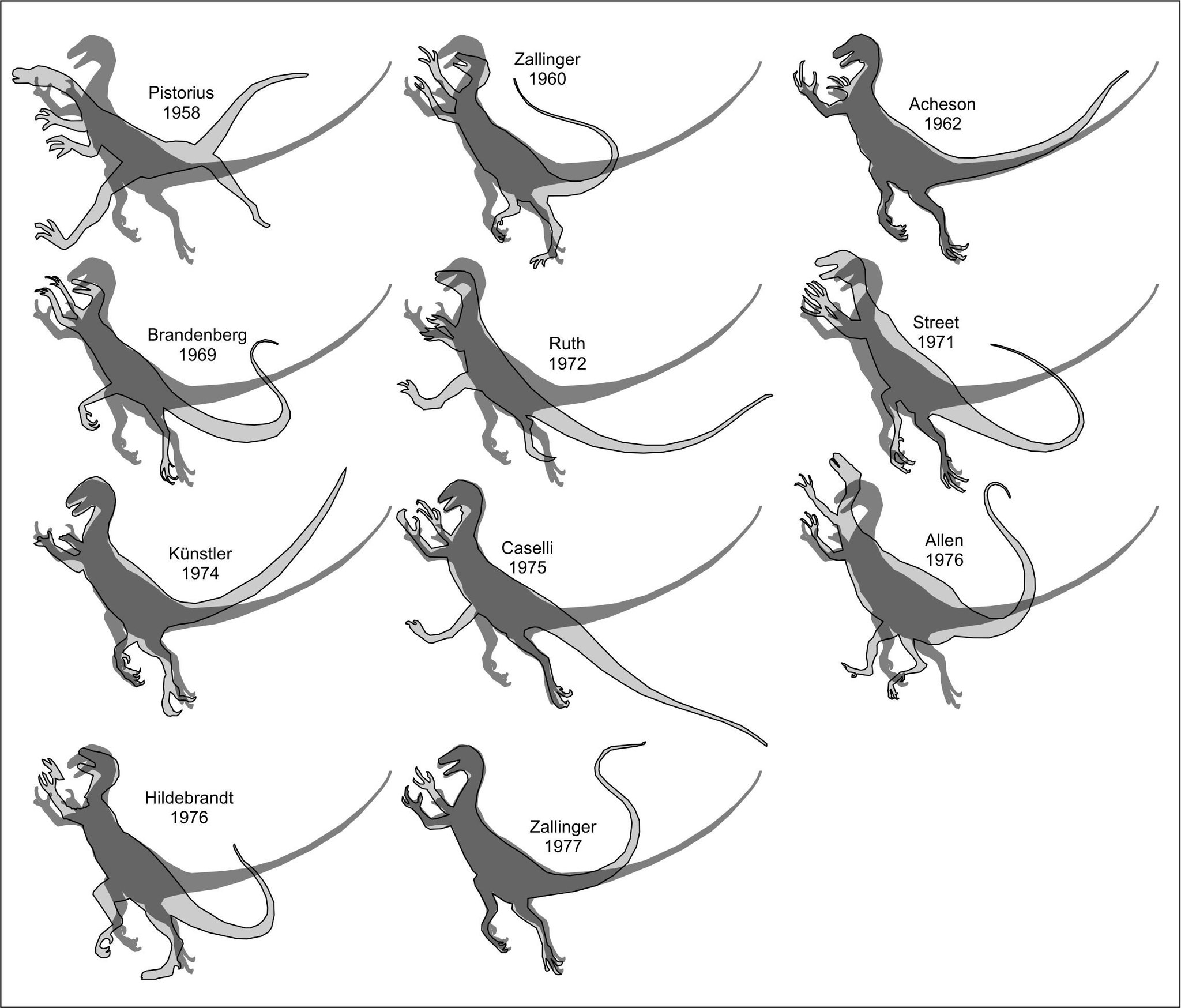
Essa mostra varie silhouette di
ricostruzioni paleoartistiche di Ornitholestes, sovrapposte
con la silhouette della prima (e abbastanza nota) ricostruzione di
Ornitholestes, realizzata nel 1914 da Charles Knight. Il nome
a fianco di ogni silhouette indica l’autore e l’anno della
ricostruzione. Notate che ci sia una qualche variabilità ma anche
una forte conservazione in determinati caratteri di queste versioni
alternative ovviamente ispirate alla immagine originaria del 1914. La
mia mente di filogenetista ha subito iniziato a ragionare se quelle
diverse silhouette fossero il frutto di un qualche processo
evolutivo, e quindi commentai sulla sua pagina Facebook di Witton che
sarebbe stato interessante fare un'analisi filogenetica di quelle
silhouette.
A quel punto, nacque l'idea del post.
Mentre ragionavo su come svolgere
l'analisi filogenetica, la presenza delle date di realizzazione
associate ad ogni immagine fece scattare una colossale deformazione
professionale: e se applicassi i metodi filogenetici Bayesiani che
includono il dato-tempo nelle analisi (oggetto della mia tesi di
dottorato)?
E così ho fatto.
Ho stilato una lista di 10 caratteri
morfologici che differenziano le varie silhouette (la lista può
essere più ampia, ma mi pare che già così sia esaustiva della
diversità nel campione):
[1] 'Mouth: open (0); closed (1).'
[2] 'Head orientation: ventral (0);
antero/dorsal (1).'
[3] 'Neck: erect (0); projected
anteriorly (1).'
[4] 'Right hand: ventral (0); at
level (1) of mouth.'
[5] 'Left arm: pointing dorsally (0);
pointing anteriorly (1).'
[6] 'Left knee: flexed (0); extended
(1).'
[7] 'Left foot: extended (0); flexed
(1).'
[8] 'Right leg: touch ground (0);
raised (1).'
[9] 'Tail base: straight (0); flexed
(1).'
[10] 'Tail tip position: level of
head (0); below head (1).'
Ho codificato tutte le silhouette
(inclusa quella di Knight 1914) per quest caratteri, ed ho caricato
la matrice corrispondente, assieme ai dati cronologici di ciascuna
silhouette in BEAST2, impostando come modello per l'albero il
“Fossilized Birth-Death” model con “Sampled Ancestors”. Si
tratta di un modello per la ricostruzione degli alberi che è in
grado di determinare se uno o più taxa analizzati siano ancestori
diretti di altri taxa inclusi nella analisi, e quindi non debbano
essere analizzati come coppie di sister-taxa divergenti ma come punti
di una singola linea evolutiva.
L'albero risultato dalla analisi è il
seguente:
 |
| L'asse tempo è quello orizzontale. Le silhouette con il nome dentro un rettangolo sono antenati diretti delle silhouette successive. |
L'analisi predice che Knight 1914 è
l'antenato di tutte le altre silhouette. Pistorius 1958 non è
antenato di alcuna silhouette successiva, e risulta sister-taxon dei
restanti discendenti di Knight 1914, che formano un clade che battezzo
Zallingeria, definito come “Zallinger 1960 e tutti i sui
discendenti”. L'analisi predice che Pistorius 1958 e Zallingeria si
sono separati memeticamente intorno al 1953. Inoltre, Zallinger 1960
genera direttamente Acheson 1962, il quale è l'antenato diretto di
tutte le silhouette successive. Intorno al 1966, la linea memetica
che porta a Branderberg 1969 si separa dalla linea che conduce agli
Ornitholestes realizzati negli anni '70, i quali formano un
clade chiamato Streetoidea, definito come “Street 1971 e tutti i
suoi discendenti”. Ruth 1972 risulta il discendente diretto di
Street 1971, ed è a sua volta l'antenato di tutte le silhouette
successive. L'analisi predice che intorno al 1973 avviene una nuova
divergenza memetica: una linea, detta Kunstleridae (definita come
“Kunstler 1974 e tutti i suoi discendenti”), si separa da
un'altra linea, detta Casellidae (definita come “Caselli 1975 e
tutti i suoi discendenti”). All'interno dei Kunstleridae, Allen
1976 è l'antenato diretto di Zallinger 1977 (che non discende quindi
direttamente da Zallinger 1960), mentre nei Casellidae, Hildebrandt
1976 è il discendente di Caselli 1975.
Interessante osservare che 2/3 delle
silhouette analizzate si possono considerare come antenati di altre
silhouette (ovvero come punti di una singola linea evolutiva), e che
la storia paleoartistica di Ornitholestes nell'intervallo
1914-1977 si possa descrivere con una singola sequenza principale e
solamente 3 eventi di divergenza “artistica” (la divergenza
Pistorius-Zallengeria, la divergenza Branderberg-Streetoidea, e la
divergenza Casellidae-Kunstleridae).
Quanto questo post è idiota e senza
senso, e quanto potrebbe essere una interpretazione plausibile delle
varie influenze che in 60 anni hanno plasmato (in modo alquanto
conservativo) l'immagine di Ornitholestes?
Lascio a voi questa valutazione.
It's a long lineage; I found another immediately after googling 'Ornitholestes'.
RispondiEliminaPaul W.
Sure, but the analysis was based on just the silhouettes in Witton's image and focuses on them.
Elimina"I found another immediately after googling 'Ornitholestes'."
RispondiEliminaThese were specifically Ornitholestes featured in the 'bird robber' trope - those images were it is running/leaping forward to catch a bird. There are a number of Ornitholestes artworks based on Knight's where the bird isn't included, but I was interested in the 'core' of the meme when putting the images together. To see most of these in more context, check out:
https://twitter.com/MarkWitton/status/828582205299494912
Also, slightly daft as this idea is, I find the results really interesting! It would be interesting to run similar analyses on other memes. I suppose the most scientific approach would be to include non-memetic depictions of the same taxon as well. 20th century Archaeopteryx pictures would be fascinating to run through this sort of assessment, although it might take a long time to code and assess. The 'giraffoid Barosaurus' might be more doable, even with non-memetic images.
I suspect a network analysis would be a more appropriate way to analyse meme evolutions as it allows hybridation and multiple ancestors for a single image.
EliminaThe interesting use of Bayesian method is that I can incorporate the age of the pictures as additional criterion for inferring relationships (assuming that pictures closer in age may have a higher probability of being linked than those with a large age distance), instead of just their morphology.
Yeah, you are right, the one I saw didn't include the bird. It seemed to be a copy of a copy with updated anatomy.
EliminaPaul W.
Scusa, ma sono più interessato a questa parte
RispondiEliminaSi tratta di un modello per la ricostruzione degli alberi che è in grado di determinare se uno o più taxa analizzati siano ancestori diretti di altri taxa inclusi nella analisi, e quindi non debbano essere analizzati come coppie di sister-taxa divergenti ma come punti di una singola linea evolutiva
Se A è l'antenato comune più recente di B e C, immagino che le coppie di sister-taxa saranno A - B e A - C. Nella visualizzazione che di solito vedo in una linea vedo prima la A al termine di una retta che si diparte della linea principale e poi successivamente B e C su altrettante rette che si dipartono dalla principale, oppure B e C su due rette che si uniscono su una sola retta che a sua volta parte dalla principale. In pratica il nodo non rappresenta nessuna specie.
Con l'impostazione che hai indicato immagino che la visualizzazione comporti un nodo (la specie A) con due rette divergenti, una per B e una per C.
In entrambi i casi A, B e c sono punti di una singola linea evolutiva.
Evidentemente mi sfugge qualcosa. Mi puoi aiutare?
No, se A è antenato di B e C, le coppie non possono essere simultaneamente A-B e AC, perché allora la topologia è una tricotomia (ABC) non risolvibile.
EliminaSe A è antenato allo stesso tempo di B e C, significa che in realtà A è una serie parafiletica di forma A1, A2, ecc..., di cui una è sister di B e una è sister di C.
Quindi, A non è un vero taxon da analizzare ma occorre spezzarlo nelle sue parti, ognuna delle quali poi può fare relazioni con B e/o C.