|
| Ricostruzione di Dakotaraptor accompagnato da due ornithuri. |
La Formazione di Hell Creek è nota
principalmente per i suoi dinosauri di taglia medio-gigante, nello
specifico i generi Tyrannosaurus,
Triceratops, Edmontosaurus e Ankylosaurus.
Tuttavia, sebbene noti da resti più frammentari ed isolati rispetto
ai generi sopra citati, nella Formazione di Hell Creek sono presenti
anche dinosauri di taglia media e piccola. Tra i theropodi, almeno
due enantiornithi e numerosi ornithuromorfi sono documentati, assieme
a resti di ornithomimidi, parvicursorini, il grande caenagnathide
Anzu, almeno un troodontide ed un numero imprecisato di
dromaeosauridi, noti principalmente per i denti. Recentemente, sempre
da questa formazione è stato istituito un nuovo dromaeosauride,
Acheroraptor, noto solamente da parte del muso. A parte
Acheroraptor, gli altri dromaeosauridi sono dedotti dai
differenti morfotipi di denti isolati rinvenuti.
Un nuovo dromaeosauridae da livelli
tardo-maastricthiani del South Dakota è descritto da De Palma et al.
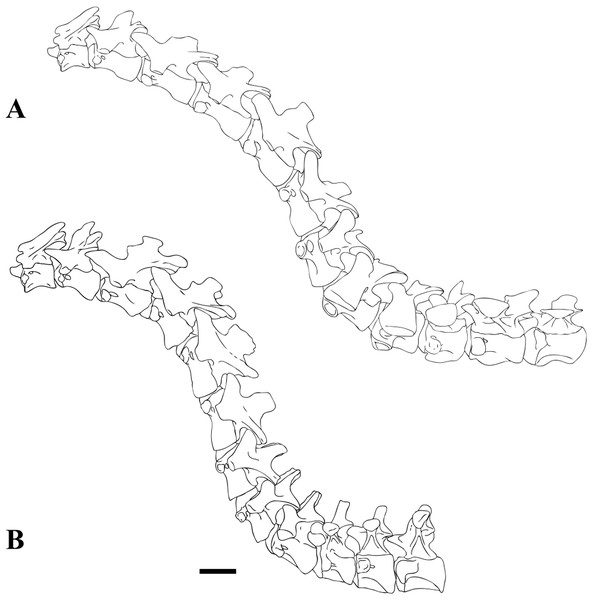
(2015): Dakotaraptor steini. Basato su resti associati che
indicano un animale lungo almeno 5 metri, e da resti riferiti ma di
dimensioni minori, Dakotaraptor è noto da alcuni denti,
alcune vertebre caudali medio-distali, la furcula, omero ed
avambraccio quasi completi, un secondo metacarpale e alcuni frammenti
di falangi, femore e tibia completi e frammenti del piede.
L'attribuzione a Dromaeosauridae è basata sulla disparità
dimensionale dei denticoli nei denti, la presenza di prezigapofisi
estremamente allungate nelle caudali intermedie, e la presenza di un
ungueale falciforme nel piede (probabilmente, del secondo dito) che
presenta il margine ventrale affilato ed i solchi collaterali posti
asimmetricamente. L'ulna presenta delle vistose papille ulnari per
l'inserzione delle penne remiganti secondarie: si tratta della prima
evidenza diretta di penne remiganti in un Dromaeosauridae di grande
dimensione, attributo finora ritenuto comunque plausibile in base
alla inferenza filogenetica (come sa bene chi legge questo blog da molti anni): da oggi, se qualcuno volesse ancora sostenere che i
dromaeosauridi di grandi dimensioni erano privi di piumaggio sarà
istigato a scrivere 100 volte alla lavagna: “l'inferenza
filogenetica è un metodo accurato di ricostruzione del tegumento nei
dinosauri”.
La presenza di un grande
Dromaeosauridae (comparabile in lunghezza ad Utahraptor e
Austroraptor, sebbene le proporzioni più slenciate delle ossa
degli arti indichino che Dakotaraptor era meno massiccio di
Utahraptor) nella formazione di Hell Creek aggiunge un
tassello alla rete trofica, che si situa in una area che prima si
riteneva essere occupata solo dai giovani di Tyrannosaurus:
dato che i tyrannosauridi di taglia medio-piccola sono
ecomorfologicamente più cursori dei dromaeosauridi, è comunque
plausibile che questi due taxa non fossero strettamente in
competizione sul piano ecologico.
Dato che questo nuovo dromaeosauridae
sta già occupando l'immaginario autoerotico di molti dinomaniaci,
esaltati dall'avere finalmente la prova che il Nord America del
Maastrichtiano terminale era niente altro che una gigantesca Isla
Nublar cinematografica con un grande dromaeosauridae assieme a T.
rex, dico subito che è totalmente ridicolo qualsiasi discorso su
una competizione diretta tra gli adulti di Dakotaraptor e
quelli di Tyrannosaurus, la enorme differenza dimensionale (il
secondo era almeno 12-15 volte più massiccio del primo), morfologica
(e quindi, ecologica) tra questi due theropodi esclude che ci fosse
una competizione diretta tra (gli adulti di) questi taxa.
DePalma et al. (2015) includono
Dakotaraptor in una analisi filogenetica, da cui risulta un
dromaeosaurino sister-group di Dromaeosaurus. Purtroppo, non
sono presentati i dettagli di questa analisi, né le sinapomorfie a
supporto di tale risultato.
Immesso in una versione ridotta di
Megamatrice, Dakotaraptor risulta sister taxon del nodo
“Dromaeosaurinae + Velociraptorinae” in quanto non presenta i
denticoli distali uncinati e presenta un ipocleido (sebbene di forma
inusuale, più ampio che lungo), ed è escluso dal nodo
“Dromaeosaurinae + Velociraptorinae” in quanto presenta centri
caudali intermedi con una cresta laterale: tutti questi caratterei
comunque hanno una discreta omoplasia (difatti l'indice Bremer dei
nodi è basso in questa area dell'albero), pertanto questo risultato è probabilmente provvisorio.

Analisi utilizzando l'implied weighting di TNT collocano Dakotaraptor in Velociraptorinae, sebbene non ci sia un buon consenso tra le differenti topologie.

Analisi utilizzando l'implied weighting di TNT collocano Dakotaraptor in Velociraptorinae, sebbene non ci sia un buon consenso tra le differenti topologie.
Da notare che nessuna di queste analisi
supporta una affinità diretta tra Dakotaraptor e l'altro
dromaeosauridae del Maastrichtiano finale nordamericano,
Acheroraptor. Basandomi sulle
foto dei denti di Dakotaraptor,
questo ultimo pare non presentare le marcate creste apicobasali nelle
corone, diagnostiche per Acheroraptor,
avvalorando una distinzione tra i due taxa. A questo proposito, è
interessante rimarcare che gli esemplari riferiti di Dakotaraptor,
pur appartenendo ad individui grandi circa 4/5 dell'olotipo, sono
riferibili ad esemplari maturi: ciò potrebbe indicare due specie di
Dakotaraptor, di
diversa taglia adulta, oppure una ampia variabilità dimensionale in
questa specie. Acheroraptor potrebbe essere il morfotipo più piccolo?
Bibliografia:
Robert A. DePalma, David A. Burnham, Larry D. Martin†,Peter L. Larson and Robert T. Bakker (2015) The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation. Paleontological Contributions 14 (16 pp.)